


Виды динамики экосистем
Любая экосистема постоянно находится в состоянии динамики (изменений), вызванной как изменением факторов среды, то есть внешними возмущениями, так и внутренними процессами. При этом удобно различать циклическую динамику и поступательную. К разряду циклической динамики можно отнести:
- суточную динамику, связанную с изменением активности растений и животных в течение суток: изменения в фотосинтезе, транспирации (испарение воды с поверхности листьев), у животных это смена периодов активности и сна и т.п.;
- сезонную динамику (смена периодов вегетации и покоя), связанную с сезонными колебаниями: смена времен года, периодов дождей и засухи, муссонная периодичность и т.п.;
- многолетнюю цикличность, вызванную, например, периодичностью солнечной активности (период порядка 10-11 лет), или же какими-то другими колебаниями с многолетним периодом, например, цикл Эль-Ниньо, который приводит к усилению ураганов, обрушивающихся на Южную Америку с периодичностью 5…7 лет, и т.п.
Особого внимания заслуживает поступательная динамика экосистем, или развитие. Обычно развитие сопровождается последовательным рядом изменений видовой и трофической структур экосистемы, всей ее организации, что позволяет даже говорить о последовательной смене экосистем, поэтому данный процесс называется сукцессией (от латинского слова сукцессио - преемственность, наследование). В зависимости от причин сукцессии различают
- экзодинамические (от греческого слова эксо - снаружи) сукцессии, вызванные внешними по отношению к данной экосистеме факторами;
- эндодинамические (от греческого слова эндон - внутри) сукцессии, вызванные внутренними механизмами экосистемы (в теории управления различают вынужденную и свободную составляющие динамического процесса, которые соответствуют двум видам сукцессии экосистем).
Экзодинамические сукцессии могут быть вызваны изменениями климата, понижением уровня грунтовых вод, подъемом уровня мирового океана и т.п. Такие смены могут длиться столетиями и тысячелетиями. Они связаны в основном с действием механизмов адаптации экосистемы к факторам внешней среды, которые в свою очередь базируются на механизмах адаптации живых организмов экосистемы.
Эндодинамические сукцессии приводятся в действие особыми законами, механизмы которых до сих пор во многом непонятны. Известно, что на любом, даже абсолютно безжизненном, субстрате типа песчаных дюн или затвердевшей лавы, рано или поздно расцветает жизнь. При этом формы жизни, точнее, типы сообществ, в данном пространстве последовательно сменяют друг друга, постепенно усложняясь и увеличивая видовое разнообразие, формируя так называемый сукцессионный ряд, состоящий из последовательных стадий, отмечающих смену одного сообщества другим. Сукцессионный ряд заканчивается стадией зрелости, на которой экосистема изменяется очень мало. Экосистемы на этой стадии называются климаксными (от греческого слова климакс - лестница).
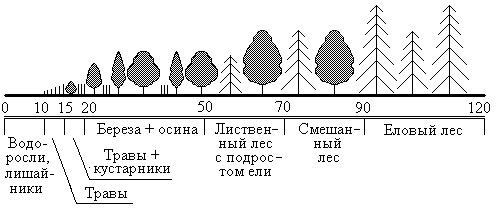
Рис.12.1
Продолжительность сукцессии от зарождения экосистемы до стадии климакса может составлять до сотен и даже до тысяч лет. Столь длительная продолжительность связана в основном с необходимостью накопления питательного вещества в субстрате. Изначально на безжизненном субстрате поселяются так называемые пионерные виды, типа лишайников и корковых водорослей (рис.12.1). За 5…10 лет они несколько обогащают субстрат питательными веществами, формируя зачатки почвы. Затем на этих еще совсем бедных почвах поселяются травы, еще более обогащая почву. Лет через 15 от начала сукцессии на когда-то безжизненном пространстве поселяются первые кустарники, которые постепенно вытесняются лиственными светолюбивыми деревьями, чаще всего березой и осиной, которые характеризуются быстрым ростом. К 50-летнему возрасту в молодом лиственном лесу выделяются наиболее сильные деревья, которые затеняют более слабую поросль, которая погибает, давая возможность поселиться под пологом лиственного леса поросли ели. Ель более теневынослива, под защитой лиственных деревьев она постепенно догоняет их в росте, отвоевывая у них жизненное пространство. Где-то к 70 годам экосистема достигает стадии смешанного елово-лиственного леса. Лиственные деревья к тому времени успевают состариться, и постепенно ель выходит на первый ярус, затеняя и изреживая всю лиственную растительность. К 90 годам данная экосистема достигает стадии климакса, для которой характерно практически полное отсутствие лиственных деревьев, ель становится доминирующим видом-эдификатором, формируя особым образом всю жизнь населяющего данную экосистему сообщества.
Это типичный пример сукцессии, характерный для южной тайги. Как ни странно, в одном и том же регионе практически все сукцессии развиваются по довольно похожим сценариям и заканчиваются в итоге одним и тем же климаксом. Конечно, в зависимости от конкретных особенностей среды возможны некоторые вариации общего сценария, но они не принципиальны. Таким образом, каждый достаточно крупный регион характеризуется своей программой развития экосистем и своим особым климаксом, который в данном смысле называется моноклимаксом. Это еще один аргумент в пользу биополевого механизма координации жизни в биосфере - каждый регион планеты развивается в соответствии с характерной для него программой и имеет свою характерную структуру, призванную обеспечить определенную функцию, закрепленную за данным регионом в едином организме биосферы (или планеты).
Правда, справедливости ради надо сказать, что у автора концепции моноклимакса, американского эколога Клементса, имеется достаточное количество оппонентов. Может быть, это связано с тем, что любая крайне полярная точка зрения всегда оказывается односторонней. Концепция моноклимакса хорошо укладывается в холистский подход, однако сторонники исследования детальных механизмов природных явлений приводят множество примеров, когда в районах с известным моноклимаксом формируются совершенно другие климаксные экосистемы. Например, в южной тайге наряду с еловыми лесами могут формироваться луговые и сосновые климаксные экосистемы. Сторонники моноклимакса склонны рассматривать их как промежуточные стадии, которые рано или поздно обязательно должны прийти к моноклимаксу, если для этого не будет таких препятствий, как, например, вмешательство человека. Так луговые экосистемы, похоже, существуют только за счет регулярного скашивания молодой древесной поросли. Сосновые леса могут возникать и без участия человека, особенно на крайне бедных почвах, типа песчаных и щебнистых почв, куда более требовательная к почвам ель внедриться не может. Однако по мере накопления плодородия почв, ель, как более сильный эдификатор, скорее всего, вытеснит сосну и создаст моноклимаксную экосистему.
Проверить это достаточно сложно, так как подобного рода сукцессии растянуты на тысячи лет. Иногда экологам улыбается удача, и они обнаруживают экосистемы, где можно непосредственно «наблюдать» такие тысячелетние сукцессии. Это позволяет выявить некоторые характерные особенности сукцессионного процесса. Так, например, замечательной природной лабораторией являются дюны у южной оконечности озера Мичиган (США, штат Индиана). Озеро постепенно отступает, оставляя после себя все более молодые песчаные дюны, абсолютно безжизненные, которые постепенно «осваиваются» жизнью. Процесс этот длится уже тысячи лет, поэтому разноудаленные от кромки воды дюны находятся на разных стадиях сукцессии, что позволяет проследить весь ход сукцессионного процесса. Первые поселенцы на дюнах - это прибрежные злаки, ива, песчаная вишня, хлопковое дерево и животные: пауки, кузнечики и т.п. За ними следуют сухие сосновые леса, затем сухие леса из черного дуба, и, наконец, на самых старых дюнах - влажные леса из дуба, бука и клена, для которых характерна мощная богатая гумусом почва, населенная дождевыми червями и улитками. Для достижения климаксного сообщества на дюнах требуется около 1000 лет, что примерно в пять раз больше, чем это требуется при более лучших, чем дюны, исходных условиях.
В зависимости от исходных условий принято различать первичные сукцессии, которые начинаются на абсолютно безжизненных субстратах, например, на дюнах, и вторичные сукцессии, начинающиеся с более благоприятных стартовых условий, например, после пожара, вырубки леса или на заброшенном поле. Вторичные сукцессии могут начинаться с более поздних пионерных видов, например, с трав или кустарников. Так, например, сукцессии на заброшенных полях начинаются с однолетних трав, бывших ранее сорняками. Затем они сменяются многолетними травами, кустарниками и деревьями. Ввиду большего исходного плодородия почвы такие сукцессии достигают климакса гораздо быстрее, чем в случаях первичной сукцессии. В последнее время для биосферы характерны именно вторичные сукцессии, что связано главным образом с деятельностью человека.
Вмешательство человека в природу ведет зачастую также к явлениям дигрессии (от латинского слова дигрессион – отклонение), когда климакс экосистемы (устойчивое равновесное состояние) достигается на более ранних стадиях сукцессии, вследствие чего экосистемы значительно упрощаются. В отличие от моноклимакса эта стадия называется катаценозом (от греческого слова ката – вниз, против). Типичным примером является пастбищная дигрессия, когда вместо леса, на данном пространстве формируется обширная луговая экосистема. Чем дальше отступает пояс лесов, тем реже достигают данного пространства семена кустарников и деревьев. Регулярный выпас скота не дает развиться молодым деревьям. Постепенно на этом месте вдали от лесов формируется особая зона, богатая гумусом, где никогда (точнее, очень долго) не вырастет лес, даже если человек оставит этот район. Так, вероятно, формировались степи, на которых древние кочевые племена пасли свои табуны и стада. Попытки возобновить леса в степной зоне требуют немалых усилий, так как помимо колонии деревьев, лес характеризуется особой структурой симбиотических связей, особенно между конями деревьев и грибницей (микроза). Поэтому лес может постепенно «наступать» на степь. Но, чтобы лес самопроизвольно возник посреди степи - случай практически невозможный. Для того, чтобы лес заменил степи после ухода человека потребуются, вероятно, десятки тысяч лет.
Следует различать автотрофные и гетеротрофные сукцессии. Все описанные выше примеры относятся к автотрофным сукцессиям, так как они протекают в экосистемах, где центральным звеном является растительность (фитоценоз). Динамика гетеротрофов целиком подчинена динамике автотрофов - смена животных сообществ зависит от смены растительных сообществ. Автотрофные сукцессии могут длиться теоретически вечно, так как они постоянно подпитываются энергией Солнца.
В гетеротрофных сукцессиях участвуют только животные (гетеротрофы, консументы). В этот процесс могут быть также вовлечены и мертвые растения, например, поваленные деревья, пни и т.п., которые являются, как правило, источником энергии для гетеротрофной сукцессии. Примером такой сукцессии может служить разрушение поваленного дерева. Первыми на мертвом или ослабленном дереве поселяются насекомые-короеды, которые повреждают кору, давая возможность проникнуть в места повреждений спорам грибов. Грибы-пионеры размягчают древесину, окрашивая ее в разные цвета. Это облегчает появление следующей волны насекомых, которые питаются уже древесиной (ксилофаги). В поврежденной ими древесине развиваются грибы-деструкторы, способствующие появлению мягкой гнили. Затем их сменяют грибы-гумификаторы, превращающие гнилую древесину в гумус. На всех стадиях гетеротрофной сукцессии также идет соответствующая смена видов микроорганизмов. В конечном итоге все органическое вещество разлагается до конечных продуктов: минеральных веществ и углекислого газа. Аналогичные процессы идут и при разложении трупов животных, а также в экскрементах животных, в лесной подстилке, в загрязненных водах и т.д.
Гетеротрофная сукцессия предполагает обязательное наличие определенного запаса энергии, аккумулированной в органическом веществе. Она заканчивается вместе с исчерпанием ресурса энергии, то есть после полного разложения исходного субстрата. После этого экосистема перестает существовать. То есть для нее не определено понятие климакса. В отличие от биогеоценозов, такие экосистемы смертны.
Гетеротрофная сукцессия хорошо ассоциируется с обществом, зависящим от ископаемого горючего. Динамика гетеротрофной сукцессии описывается кривой с быстрым ростом числа организмов, пока не будет достигнут некоторый максимум, затем количество организмов постепенно уменьшается по мере исчерпания энергетического ресурса (рис.12.2).
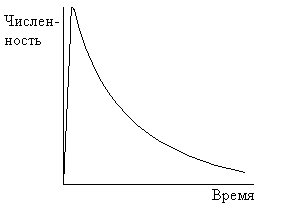
Рис.12.2
Какого-то устойчивого состояния (климакса) добиться не удается. Такое общество быстро прогрессирует, но, тем не менее, оно изначально обречено на вымирание. Мы уже «сняли сливки» с большинства месторождений. Дальнейшая их эксплуатация со временем будет требовать все больших вложений энергии. Поэтому эффективность добычи полезных ископаемых будет неуклонно падать. Вместе с этим будет падать и жизнеспособность цивилизации, построенной на гетеротрофной сукцессии, если, конечно, еще раньше не произойдет катастрофических изменений, например, массового психоза - людям свойственно искать виновников своих проблем. Поэтому мы огромное количество усилий направляем на поиск новых источников энергии. Но даже если мы научимся управлять термоядерным синтезом, это не изменит нашей деструктивной сущности. Мы только несколько отодвинем в будущее развязку, как это мы делали уже не раз, но стабилизировать гетеротрофную сукцессию нам не удастся. Это даже если не учитывать, что в случае успеха в области энергетики перегрев биосферы неизбежен.